Es ist ein besonderes Erlebnis, die weiß blühende Form einer Orchidee zu finden, die sonst üblicherweise in einer anderen Farbe blüht.
Zunächst ein Ergebnis genetischer Mutationen, sind diese Sonderformen bei manchen Orchideenarten regelmäßig anzutreffen, auch wenn sie insgesamt eher selten auftreten. Sie belegen die lebendige und vielfältige Entwicklung dieser Pflanzenfamilie.
In der botanischen Taxonomie werden sie als Varietät oder als Form betrachtet. Gelegentlich wurden sie somit auch als var. albiflora oder f. albiflora wissenschaftlich beschrieben, etwa Orchis quadripunctata f. albiflora.
Warum können wir in der Natur Orchideen ein und derselben Art mit unterschiedlichen Farben finden? Die Antwort hängt vom genetischen Bauplan der jeweiligen Art ab.
1. Einige Orchideen haben ein genetisches Modell, das Blüten in verschiedenen Farben ermöglicht. So hat zum Beispiel Dactylorhiza sambucina meistens gelbe Blüten, aber es gibt auch oft Pflanzen mit roten Blüten. Gelb blühende Pflanzen bringen gelb blühende Nachkommen hervor und umgekehrt. Aber bei Kreuzungen zwischen beiden sind auch Zwischenfarben entstanden (Svante Malmgren/Henric Nyström). Dieses Nebeneinander von zwei Farben scheint ein besonderes Kennzeichen von Dactylorhiza sambucina zu sein.
2. Farbänderungen sind eine von mehreren Möglichkeiten für die Entwicklung abweichender Varianten einer Orchideenart. Arten mit einer großen Formenvielfalt der Blüten haben oft auch unterschiedliche Blütenfarben – z.B. Anacamptis morio:

3. Jenseits dieses breiten Spektrums von Variationen zeigen einige Orchideen auch individuelle genetische Merkmale, die zu weißen Blüten führen. Diese Formen werden im allgemeinen als genetische Abweichungen verstanden, als Abwesenheit derjenigen Pigmente, welche die Farbe einer Blüte bestimmen. Die damit verbundenen Begriffe lauten Hypochromie (als Gegenbegriff zur Hyperthermie, die eine exzessive Pigmentierung, also eine besonders intensive Färbung, bezeichnet) oder Albiflora-Formen. Als alba oder albiflora bezeichnete Formen haben Blüten, denen jede farbige Pigmentierung fehlt und die rein weiß sind (Olaf Gruss: Albino Forms of the Slipper Orchids. In: Orchid Digest. Vol 69 (4), 2005. S.204). Die Bildung roter Blütenfarbstoffe kann bei einzelnen Individuen gelegentlich gehemmt oder völlig blockiert sein, so dass bei Arten, die nur rote Blütenfarbstoffe besitzen, dann blassrote bis weiße Farbformen entstehen (Wolfgang Wucherpfennig: Die Orchidee des Jahres 2007: das Schwarze Kohlröschen Nigritella nigra subsp. rhellicani, ein Kleinod der Berge. In: Berichte aus den Arbeitskreisen Heimische Orchideen Jg. 24 - Heft 1 - 2007. S.26). Ein Beispiel ist die weiße Variante von Anacamptis pyramidalis.

4. Manche bezeichnen bereits diese individuelle genetische Besonderheit als Albinismus: In diesem Verständnis wird eine Pflanze mit Blüten ohne rote Pigmente umgangssprachlich als Albino bezeichnet - die Blüten können, hellgrün, gelblich oder weiß sein. Aber in einem strikten botanischen Sinn ist ein Albino eine Pflanze, der Chlorophyll fehlt: Daher ist Albinismus die völlige Abwesenheit von grünem Pigment. Da die meisten hier vorgestellten Orchideen grüne Blätter haben, handelt es sich also nicht um Albinos, sondern um Albiflora-Formen. Manchmal herrscht die weiße Farbe nur in einem Teil der Blüte vor. In anderen Fällen ist die ganze Blüte reinweiß, einschließlich der Pollinien.
Nur wenn die ganze Pflanze völlig weiß ist, einschließlich der Blätter, kann man von einem Albino sprechen. Dies ist etwa beschrieben worden für Epipactis helleborine oder für Malaxis bayardii im US-Staat Massachusetts, benannt als Malaxis bayardii f. kelloggiae (Paul Martin Brown: An Albino Adder's Mouth from Cape Cod, Massachusetts. In: North American Native Orchid Journal, 11/2005. p. 4-5).
Sowohl Albinismus als auch Albiflora-Formen werden genetisch rezessiv vererbt - ein Abkömmling kann die weißen Blüten behalten, kann aber auch Blüten in der üblichen Farbe der Art entwickeln. Bei einigen Arten sind die weiß blühenden Formen relativ häufig und bilden stabile Bestände aus. Bei tropischen Orchideen haben Albino-Formen ihren eigenen floristischen Wert und werden daher entsprechend gezüchtet. Dieses Interesse hat schon früh Forschungsarbeiten angeregt, die zu interessanten Ergebnissen geführt haben. So war es zum Beispiel möglich, durch das Kreuzen von zwei weißen Orchideen eine farbige Orchidee zu erzeugen.
Begriffe wie "Abweichung", "Anomalie" oder "Mangel" beziehen sich auf eine bestimmte Norm oder einen Standard und definieren die weiß blühende Orchidee nur aufgrund des negativen Sachverhalts, nicht dieser Norm zu entsprechen. Diese Begrifflichkeit mit ihren abwertenden Assoziationen soll hier nicht weitergeführt werden. Statt dessen werden weiß blühende Orchideen einer farbig blühenden Art als Besonderheit betrachtet, mit der die Natur einen neuen Weg einschlägt. Das Ergebnis wird man erst in einigen Millionen Jahren kennen.

Orchis simia f. albiflora (links) und Orchis purpurea x simia (rechts)
Genetik
Der erste Botaniker, von dem bekannt ist, dass er zwei verschiedene Pflanzenarten gekreuzt hat, war der britische Gärtner Thomas Fairchild (1667-1729). Er kreuzte zwei Nelkenarten, indem er den Pollen von Dianthus barbatus auf den Stempel von Dianthus caryophyllus brachte, um so eine Hybridpflanze zu erzeugen. Fairchild wurde damals sehr kritisiert, man warf ihm vor, Gottes Schöpfung zu manipulieren. Aber das Interesse an neuen Arten von gärtnerisch reizvollen Blütenpflanzen war größer.
Lange Zeit wurde angenommen, dass das Kreuzen von Pflanzen mit verschiedenen Blütenfarben so funktioniert wie die Mischpalette eines Künstlers. Ein Hybrid aus einer rot und einer weiß blühenden Pflanze müsste somit rosa blühen. 1866 wies der Augustiner-Mönch Gregor Johann Mendel nach, dass dies falsch ist. Mendel (1822-1884) entdeckte das Konzept der rezessiven Allelen, eine der Erkenntnisse, die später als Mendel'sche Vererbungsgesetze bezeichnet wurden. Eine genetische Allele (oder DNA-Sequenz) ist rezessiv, wenn das Individuum einer bestimmten Art zwei Kopien des zuständigen Gens benötigt, um ein bestimmtes genetisches Merkmal zum Ausdruck zu bringen. Wenn das Individuum nur eine Kopie mitbekommen hat, von der männlichen oder weiblichen Seite, bleibt dieses Merkmal verborgen - im Unterschied zu einer dominanten Allele.
Mendel studierte die Blütenfarbe von Erbsen und fand heraus, dass Purpur die dominante Allele ist und dass Weiß rezessiv ist. Individuen mit zwei Purpur-Allelen (BB) haben eine purpurne Blütenfarbe - ebenso wie Pflanzen mit einer purpurnen und einer weißen Allele (Bb). Nur Erbsen, bei denen beide Allelen weiß sind (bb) entwickeln weiße Blüten. 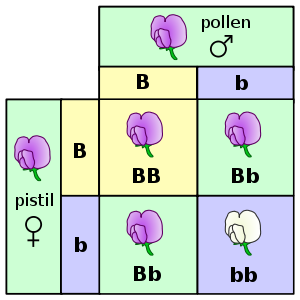
In Anlehnung an den lange vergessenen Mendel war Charles Chamberlain Hurst (1870-1947) der erste, der Albinismus bei Orchideen untersuchte. Er entdeckte, dass tatsächlich zwei Gene für die Ausbildung einer bestimmten Blütenfarbe verantwortlich sind: Faktor C ermöglicht die Bildung des Farbstoffs, während der andere Faktor, R, bestimmt, welche besondere Farbe erscheint (Rohrl, Helmut: For Heaven‘s Sake, It‘s Xanthic! Albinism in Orchids. In: Orchid Digest. Vol 69 (4), 2005. S.241). Jedes dieser Gene gibt es auch in einer inaktiven Form, c and r. Farbig blühende Pflanzen haben ein oder zwei aktive Allelen geerbt: CC und RR, Cc und RR, Cc und Rr oder CC und Rr. Albinos haben entweder die Kombination cc oder rr. C und R sind die dominanten Allelen, die die besonderen Enzyme bestimmen, die zur Produktion von Farbpigmenten erforderlich sind - unter ihnen die Anthocyanine, die für die Blütenfarbe besonders wichtig sind.
Seitdem gab es sehr viel genauere Forschungen zu den genetischen Prozessen, welche die Blütenfarbe bestimmen. Es sind mindestens 35 Gene bekannt, welche bei der Blütenfarbe von Petunien eine Rolle spielen (Timothy A. Holton/Edwina C. Cornish: Genetics and Biochemistry of Anthocyanin Biosynthesis. In: The Plant Cell, Vol. 7,1995, S. 1071). Unter ihnen sind auch Regulierungsgene, welche den Zeitpunkt, die Verteilung und die Menge der Anthocyanin-Pigmentierung beeinflussen).
Ein einzelnes Gen, also eine bestimmte Region des Chromosoms, enthält die Kodierung für die Herstellung von Enzymen, welche für die Biosynthese von Pigmenten, für ihren „biochemischen Pfad“, erforderlich sind. Die genetische Blaupause, bezeichnet als Genotyp, bestimmt so den Phänotyp, die visuellen Merkmale einer Blüte.
Forscher erzeugen transgene Pflanzen, um für die Pigmenterzeugung relevante Gene einzuführen, die natürlich nicht vorhanden sind. So gibt es etwa transgene Petunien mit Genen fremder Arten, welche die Produktion von Pelargonidin ermöglichen, einem Anthocyanin mit einer tiefroten Färbung. Es gibt zwei verschiedene Wege für eine solche gentechnische Veränderung mit dem Ziel, die Blütenfarbe zu verändern: (1) die Einführung von Genen, welche auf neue Enzym-Aktivitäten kodiert sind, und (2) die Inaktivierung von endogenen Genen (Holton/Cornish 1995, S.1077). Dies geschieht etwa durch das Verfahren der RNA-Interferenz (RNAi), bei dem gezielt einzelne Gene ausgeschaltet werden, so dass deren spezifische Eiweiß-Botenstoffe nicht mehr erzeugt werden.
Im Fall unserer Albiflora-Orchideen geht die Natur den zweiten Weg: Die Biosynthese der Blütenfarben-Pigmente ist blockiert. Da an diesem komplexen Prozess so viele verschiedene Gene beteiligt sind, gibt es auch verschiedene genetische Möglichkeiten, wie dieser Prozess blockiert oder reduziert wird.
Pigmente
Es gibt vier Gruppen von Pigmenten, die der Pflanze nicht nur verschiedene Farben bereitstellen, sondern auch unterschiedliche Funktionen für die Pflanzenbiologie erfüllen:
1) Chlorophyll: Dieses grüne Pigment ist die entscheidende Voraussetzung für die Photosynthese, die Licht und Kohlendioxid (CO2) in Glucose und Sauerstoff (O2) umwandelt. Chlorophyll nutzt die blauen und roten Bereiche der Wellenlänge des Lichts, kann die grünen Bereiche aber schlechter verwerten: Rotes und blaues Licht wird absorbiert, grünes Licht wird reflektiert. Daher sind diejenigen Teile der Pflanze, die Chlorophyll enthalten, grün. Chemisch bestehen die Pigmentmoleküle aus einem Magnesium-Kern mit vier umgebenden Stickstoff-Atomen sowie fünf bis sechs Teilen Sauerstoff, 28 bis 72 Teilen Wasserstoff und 35 bis 55 Teilen Kohlenstoff. Sie sind nicht wasserlöslich, sondern fettlöslich und kommen in der Zelle in den Chloroplasten vor. Der Name kommt aus dem Griechischen: χλωρός (chloros ist „grün“) und φύλλον (phyllon ist das „Blatt“).
2) Flavonoide: Diese wasserlöslichen Pigmente haben als Ausgangsstoff die Aminosäure Phenylalanin, die in höheren Konzentrationen etwa in der Muttermilch von Säugetieren zu finden ist. Es gibt verschiedene Gruppen von Flavonoiden.
2 a) Anthocyanine: Diese Pigmente stellen ein breites Farbspektrum bereit von orange/rot bis violett/blau. Die genaue Farbe wird mitbeeinflusst von anderen Pigmenten, Metall-Ionen und dem pH-Wert (das ändert sich von rot in einer säurehaltigen Umgebung zu blau in basenreicher Umgebung) (Yoshikazu Tanaka/Nobuhiro Sasaki/Akemi Ohmiya: Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids. In: The Plant Journal, 54/2008 , S. 733-749). Anthocyanine kommen besonders in den Blütenblättern vor - sie fehlen bei allen Albiflora-Varietäten von Orchideen. Die von den Anthocyaninen bereitgestellte Farbe hat mehrere biologische Funktionen. Die Pigmentbereiche reflektieren Licht zu Chlorophyll-haltigen Teilen der Pflanze, um so die Produktion von Glucose zu erhöhen. Darüberhinaus schützen die Anthocyanine die sensiblen Teile der Pflanze vor möglicherweise schädlicher Lichtstrahlung, indem sie blau-grünes Licht und die UV-Strahlung absorbieren. Schließlich lockt die Blütenfarbe bestäubende Insekten an. Es gibt wahrscheinlich mehr als 550 verschiedene Anthocyanine. Dazu gehören die ziegelroten Pelargonidin-, die roten Cyanidin- und die blauen Delphinidin-Pigmente.
In einem komplexen Prozess der Biosynthese werden mehr als fünf Enzyme benötigt, um die wasserlöslichen Anthocyanine in den Vakuolen der Zelle zu produzieren. Jede, auch die kleinste Störung im Mechanismus eines dieser Enzyme durch genetische Faktoren oder Umweltbedingungen kann die Produktion von Anthocyaninen stoppen.(Wikipedia:Anthocyanins). Der Name kommt aus dem griechischen ἀνθός (anthos ist die "Blüte") und κυανός (kyanos bedeutet "blau").
2 b) Flavone und Flavonole. Diese Flavonoide werden als Ko-Pigmente bezeichnet, weil sie für das menschliche Auge farblos sind, aber die Farbe von Anthocyaninen beeinflussen können. Der Unterschied zwischen beiden besteht darin, dass Flavonole eine zusätzliche Wasserstoff-Sauerstoff-Verbindung in ihrer Molekularstruktur haben. Da sie UV-Licht absorbieren, das von Insekten erkannt wird, geben sie den Blüten Farbe und Muster, um Insekten anzulocken (Tanaka/Sasaki/Ohmiya 2008, S.737). Flavone und Flavonole sind meist in weißen Blütenblättern enthalten. Es gibt bei Pflanzen keine weißen Pigmente, aber weiße Blüten reflektieren das gesamte sichtbare Licht und erscheinen daher weiß. Farblose Flavonoide geben weißen oder cremefarbenen Blüten 'Tiefe' (Erich Grotewold: The Genetics and Biochemistry of Floral Pigments. In: Annual Review of Plant Biology 2006, S.770). Die Molekularstruktur der Flavonoide beruht auf Sauerstoff und Wasserstoff-Sauerstoff-Verbindungen. Flavonole haben eine zusätzliche Wasserstoff-Sauerstoff-Verbindung.
3) Carotinoide: Diese Pigmente, die zur Gruppe der Terpenoide gehören, decken Wellenlängen von Gelb bis Rot ab. Zusammen mit bestimmten roten oder violetten Anthocyaninen ermöglichen sie braune oder bronzefarbene Farbtöne (Grotewold 2006, S.766). Carotinoide können in allen Teilen einer Pflanze vorkommen, sind aber oft von Chlorophyll verdeckt. Wie die Chlorophylle sind sie fettlöslich, ihre Behälter werden als Chromoplasten bezeichnet. Ähnlich wie die Anthocyanine unterstützen sie die Photosynthese und dienen als Schutzschirm gegen schädliche Lichtwellen. Es gibt mehr als 600 Carotenoide, deren chemische Struktur ziemlich komplex ist. Der Zerfall bestimmter Carotinoide hat auch eine wichtige Rolle bei der Produktion von Blütenduftstoffen. Der Name kommt aus dem griechischen καρότον (karoton) und dem lateinischen carota (Karotte).
4) Betalaine: Es gibt zwei verschiedene Gruppen, Die Betacyanine mit Pigmentfarben von Rot bis Violet und die Betaxanthine, die Farben von Gelb bis Orange ermöglichen. Diese Pigmente sind ähnliche wie die Anthocyanine aufgebaut, da sie wie diese wasserlösliche Komponenten in den Vakuolen sind. Sie enthalten aber anders als diese Stickstoff. Betalaine kommen nur in wenigen Pflanzenfamilien vor, Orchideen gehören nicht dazu. Betalaine und Anthocyanine schließen sich gegenseitig aus (Grotewold 2006, S.765). Der Name ist abgeleitet von der Roten Beete (Beta vulgaris) mit ihren tiefrot gefärbten Wurzeln.
Taxonomie
Wenn sich der genetische Bauplan einer Art so ändert, dass eine Pflanze weiße Blüten entwickelt, stellt dies keine hinreichende Bedingung dar, um daraus eine neue Art abzuleiten. Selbst die Einstufung als Unterart (Subspezies) ist kaum angemessen. Bereits 1905 hat ein Orchideenführer in den USA festgestellt: Bei nahezu jeder Art gibt es Albino-Launen (englisch: "albino freaks"), und das allein würde nicht ausreichen, zwei sich sonst sehr ähnlich sehende Pflanzen in zwei Arten aufzuteilen (Leeming Jelliffe, Helena Dewey/Gibson, William Hamilton: Our native orchids. New York; Doubleday, 1905, p.37).
 In taxonomischer Hinsicht, also bei der wissenschaftlichen Benennung einer Art, ist die Blütenfarbe irrelevant. Dies hat auch historische Gründe: Zu den Anforderungen für die wissenschaftlich korrekte Beschreibung einer Art gehörte auch ein Herbarbeleg, also die gepressten und getrockneten Pflanzenteile. In einem Herbarium bleibt die Farbe der Blüten aber nicht erhalten.
In taxonomischer Hinsicht, also bei der wissenschaftlichen Benennung einer Art, ist die Blütenfarbe irrelevant. Dies hat auch historische Gründe: Zu den Anforderungen für die wissenschaftlich korrekte Beschreibung einer Art gehörte auch ein Herbarbeleg, also die gepressten und getrockneten Pflanzenteile. In einem Herbarium bleibt die Farbe der Blüten aber nicht erhalten.
Die weiß blühenden Pflanzen einer Art mit einer ansonsten anderen Blütenfarbe werden als Varietät oder Form charakterisiert, kenntlich gemacht mit dem Zusatz var. oder f., z.B. in var. albiflora. Das Verfahren zur Beschreibung einer solchen Varietät oder Form ist das  gleiche wie bei der Beschreibung einer neuen Art. Nur die Taxa, bei deren Beschreibung alle Anforderungen erfüllt sind, gelten als wissenschaftlich gültig. Dazu gehört auch eine Beschreibung auf Latein. Hier als Beispiel die Beschreibung von Orchis quadripunctata var. albiflora, eingetragen von Chryssoula und Antoine Alibertis in L‘Orchidophile 87/1989, S.112.
gleiche wie bei der Beschreibung einer neuen Art. Nur die Taxa, bei deren Beschreibung alle Anforderungen erfüllt sind, gelten als wissenschaftlich gültig. Dazu gehört auch eine Beschreibung auf Latein. Hier als Beispiel die Beschreibung von Orchis quadripunctata var. albiflora, eingetragen von Chryssoula und Antoine Alibertis in L‘Orchidophile 87/1989, S.112.
Da es in vielen Fällen keine solche offizielle Bezeichnung gibt, können die weißen Formen einer Art auch vereinfacht als Form bezeichnet werden: Anacamptis morio f. albiflora.
Bezeichnungen für weiß blühende und ähnliche Formen:
- albiflorum/albiflora - weiß blühend
- album/alba - weiß
- alboflavum/alboflava - gelbweiß
- alboviride - grünlich weiß
- candidum/candida - rein weiß
- flavescens - gelblich
- virescens - grünlich
- pallidum/pallida - blass
- immaculatum/immaculata - ungefleckt
In einem Beitrag für die Berichte aus den Arbeitskreisen Heimische Orchideen (Peter Zschunke: Von "alba" bis "viridiflora" - zur Taxonomie und Biologie hypochromer Orchideen. In: Berichte aus den Arbeitskreisen Heimischer Orchideen 38 (1), 2021. S. 78-99) bin ich der Frage nachgegangen, wie albiflora-Formen von Orchideen wissenschaftlich sinnvoll bezeichnet werden sollten. In der Vergangenheit waren Botaniker ziemlich kreativ darin, solche Pflanzen zu beschreiben, mit ganz verschiedenen Namen und auf unterschiedlichen Ebenen - bis hinauf zur Subspezies. Dabei erfordert "eine individuelle genetische Beeinträchtigung, wie sie der Ausfall einer Pigmentbildung darstellt, keine Beschreibung eines Taxons", wie ich in dem Beitrag erkläre. Stattdessen schlage ich vor, "alle Individuen von Orchideen-Arten mit einer ausgeprägten genetischen Störung der Pigment-Bildung auch ohne eine förmliche Beschreibung als albiflora-Formen zu bezeichnen, mit dem meistverwendeten Begriff bei bisherigen Beschreibungen und mit einem Formbegriff in einem allgemeinen Verständnis, der als 'nomen nudum' keinen Typusbeleg verlangt".
In einem Beitrag über das taxonomische Durcheinander bei den Orchideen in Europa betont Richard Bateman, wie wichtig es ist, statt individualistischer Schlüsse eindeutige Kriterien für die Klassifizierung zu entwickeln. Der Artikel "Über die entscheidende Bedeutung einer Maximierung von Befunden und Minimierung autoritärer Spekulation zur abstammungsgeschichtlichen Klassifizierung der europäischen Orchideen" (In: Journal Europäischer Orchideen, Vol. 41/2, Juli 2009) erklärt, wie aus der Phylogenese (Evolution) von Arten eine monophyletische Klassifizierung abgeleitet werden kann (also eine Klassifizierung mit einer Gruppe von Organismen, die ein Kladogramm mit einem gemeinsamen Vorfahren bilden und alle Abkömmlinge dieses Vorfahren umfassen).
Bateman erkennt ausdrücklich den Wert morphologischer Merkmale an, betont aber, dass molekularbiologische Daten die wichtigste Grundlage für eine wissenschaftliche Taxonomie bilden sollten. Er kritisiert, dass andere Klassifizierungen oft nur "das Ergebnis persönlicher Meinung" sind und sagt: "Die Daten des 21. Jahrhunderts werden eingeschränkt, indem man freiwillig einen Ansatz der biologischen Klassifizierung beibehält, der aus dem 18. Jahrhundert stammt." Mit Blick auf taxonomische Debatten über die Gattungen Dactylorhiza/Coeloglossum, Neottia/Listera oder Gymnadenia/Nigritella klagt er: "Die Autoren entscheiden aus der Vielzahl von bereits verfügbaren Klassifikationen , welche Namen sie akzeptieren und welche sie ablehnen - so als ob sie Produkte aus den Regalen eines Supermarkts auswählen."
Beim Eintreten für klare Regeln auf der Grundlage von Erkenntnissen molekularbiologischer Forschung erwähnt Bateman auch "die Entstehung von weiß blühenden Individuen bei vielen Orchideenlinien" als ein Ergebnis von konvergenter Evolution - dieser Begriff beschreibt im Allgemeinen die Entwicklung ähnlicher Merkmale bei Arten, die genetisch nicht miteinander verwandt sind, oft angestoßen durch eine Anpassung an Umweltbedingungen oder eine bestimmte funktionale Entwicklung. Aber gibt es wirklich eine solche Anpassung, wenn Orchideen weiße Blüten entwickeln, "was durch die Unterdrückung eines beliebigen Abschnitts im biosynthetischen Pfad für die Erzeugung von Anthocyanin-Pigmenten erreicht wird"? Und was wäre das Ziel einer solchen Anpassung?
Bestäuber
Während das menschliche Auge aus einer einzigen Linse besteht, sehen Insekten ihre Umwelt mit einem Facettenauge aus mehreren tausend Ommatidien (Einzelaugen).  Jedes einzelne hat einen eigenen Satz von Photorezeptoren, um visuelle Signale aufzunehmen: Bei Bienen sind das vermutlich sechs Grün-Rezeptoren, die für die Erkennung von Bewegungen zuständig sind, sowie ein oder zwei Rezeptoren für UV-Strahlung und Blau (Lars Chittka/Nigel E. Raine: Recognition of Flowers by Pollinators. In: Current Opinion in Plant Biology 2006, S.429).
Jedes einzelne hat einen eigenen Satz von Photorezeptoren, um visuelle Signale aufzunehmen: Bei Bienen sind das vermutlich sechs Grün-Rezeptoren, die für die Erkennung von Bewegungen zuständig sind, sowie ein oder zwei Rezeptoren für UV-Strahlung und Blau (Lars Chittka/Nigel E. Raine: Recognition of Flowers by Pollinators. In: Current Opinion in Plant Biology 2006, S.429).
Andere Insekten wie die Schwalbenschwanzart Papilio glaucus haben hingegen auch Rezeptoren für Rot, wie das Farbsehvermögen von Schmetterlingen wohl generell größer ist als das der Bienen (Adriana Briscoe: What colors do insects see? In: Orchid Digest 2005, S.265). Die Auflösung des Facettenauges ist etwa 100 Mal schlechter als die des menschlichen Auges. Bienen sind ziemlich kurzsichtig: Eine Blüte muss eine riesige Größe von 26 Zentimetern haben, um aus einem Meter Entfernung erkannt zu werden; um eine Blüte mit einem Durchmesser von einem Zentimeter zu sehen, muss eine Biene die Distanz auf 11,5 Zentimeter verkürzen (Chittka/Raine 2006, S.428f.).
Viel besser als das Sehvermögen ist die Fähigkeit von Insekten, Gerüche wahrzunehmen, da Bienen in ihren Fühlern mindestens 130 Rezeptoren dafür haben. Es wäre somit eine sinnvolle Strategie der Evolution, riesige Blüten zu entwickeln, um Befruchter anzulocken. Aber größere Blüten sind in der Entwicklung mit höherem Energieaufwand verbunden, und die Blütengröße wird von einer komplexen Abfolge von Interaktionen zwischen Genen und Prozessen eingeengt, die die Organentwicklung regeln (Heather M. Whitney/Beverley J. Glover: Morphology and development of floral features recognised by pollinators. In: Anthropod-Plant Iteractions 2007, S.148). Die Form der Blüte ist variabler, aber nicht so sehr wie die Farbe, welche von einem komplexen Zusammenspiel von Genen, Enzymen, Metallionen und pH-Wert bestimmt wird. Experimente zeigen, dass Hummeln und Taubenschwänzchen die Farbe von Blütenblättern deutlich unterscheiden. Diese Ergebnisse machen es wahrscheinlich, wenn auch nicht sicher, dass die Farbe der einzige wesentliche Faktor ist, wenn Befruchter die Wahl treffen (Whitney/Glover 2007, S.153). Offenbar lernen Insekten die Farben und Formen von Blüten, die sie mit Nektar belohnt haben. Da viele Orchideen keinen Nektar erzeugen, könnte es für sie lebenswichtig sein, dass sie mit einer Pflanze verwechselt werden, welche ihren Besucher mit Nektar entlohnt.
Aber eine erfolgreiche Befruchtung hängt von vielen Faktoren (ab), die außerhalb der Kontrolle einer einzelnen Pflanze oder Art sind (Chittka/Raine 2007, S.433).
Ein entscheidender Faktor ist die Nachbarschaft von anderen Blüten. Da weiße, UV-absorbierende ... Blüten in praktisch allen gemäßigten Zonen Europas und des Mittelmeerraums am häufigsten vorkommen (Chittka/Raine 2007, S.433), könnte es eine gute Strategie sein, weiße Blüten zu entwickeln: Eine Orchidee mit einer häufigen Farbe kann ziemlich sicher sein, sich in der Nachbarschaft von anderen Blüten mit einer ähnlichen Farbe zu befinden (Chittka/Raine 2007, S.434).
Zusätzlich zur Farbe beeinflusst das Muster von Blütenblättern die Art, wie Insekten Blüten erkennen. Die Linien und Flecken auf Petalen und Sepalen erzeugen einen "Flimmereffekt" aufgrund der Lichtintensität, die sich ändert, wenn ein Insekt über eine Blüte mit einem markanten Muster fliegt, und bei einem Experiment bevorzugten die Bienen diejenigen (Blüten) mit dem größten Flimmereffekt (Harold Koopowitz: More on Insect Vision in Flowers. In: Orchid Digest 69/2005. S.267). Bei manchen Albiflora-Varietäten von Orchideen bleibt die Farbe im Blütenmuster erhalten, während die umgebenden Flächen alle ihre Pigmente eingebüßt haben. Aber auch völlig weiße Blüten können ihr eigenes Muster haben, erzeugt von den UV-absorbierenden Flavonolen in den Blütenzellen: Blüten, die von Insekten mit UV-empfindlichen Augen besucht werden, haben oft Muster oder Markierungen, die von den Insekten bemerkt werden, aber die für Menschen unsichtbar sind (Koopowitz 2005, S.268).
Bienen und andere Insekten suchen Blüten auf, um Nahrung zu finden wie Nektar oder Pollen. Und sie lernen, eine Vielzahl von Blütenmerkmalen mit solchen Belohnungen zu verbinden, wie drei Forscher an der Universität von Arizona bestätigt haben: Avery L. Russell, China Rae Newman und Daniel R. Papas haben dazu einen Aufsatz in der Zeitschrift Evolutionary Ecology (DOI 10.1007/s10682-016-9848-1) veröffentlicht.

Andrena spec. mit Orchis simia
Das Arizona-Projekt interessiert sich für Pflanzenarten mit Blüten in verschiedenen Farben, aufgrund einer Mutation, bei der die Produktion von Blütenpigmenten blockiert wird. Ähnlich wie bei den auf dieser Web-Site gezeigten Albiflora-Orchideen können solche Farbe-Polymorphismen auch in anderen Pflanzenfamilien auftreten. Die Autoren erklären, dass diese recht häufig auftreten und nennen als Beispiele Geranium thunbergii, Antennaria dioica (Katzenpfötchen) oder Aquilegia coerulea (Rocky-Mountains-Akelei), eine Verwandte der europäischen Akelei.
Ihre Studie analysiert das Verhalten von Bestäubern bei einem purpurfarbenen und weiß blühenden Nachtschattengewächs (Solanum tridynamum), wobei sie eine Experimentalumgebung konstruierten. Damit konnte gezeigt werden, dass anfänglich naive Bienen ohne Lernerfahrung keine Präferenz für purpurn oder weiß blühende Pflanzen hatten. Dies war auch der Fall, als die Forscher die Entnahme von Pollen verhinderten - indem sie den Staubbeutel mit Klebstoff verschlossen. Aber Bienen mit einer lohnenden Erfahrung bei purpurnen Blüten zeigten eine signifikant starke Präferenz, auf Morphen mit purpurnen Blütenblättern zu landen, verglichen zu Morphen mit weißen Blütenblättern. jedoch war diese Präferenz für eine lohnende Farbe bei weißen Blüten mit einer lohnenden Erfahrung viel schwächer aussgeprägt. Die Autoren kommen so zur Schlussfolgerung: Bienen haben eine einseitige Neigung in der Frage, wie Erfahrung Präferenzen formt: Die Erfahrung mit dem purpurnen Morph hatte eine größere Wirkung als die Erfahrung mit dem weißen Morph.Der Grund für diese einseitige Lernerfahrung könnte nach Einschätzung der Forscher aus Arizona darin liegen, dass purpurne Blüten einen viel größeren Farbkontrast zur Umgebung aufweisen. Daher ist das hypochrome Solanum tridynamum bei der Nahrungssuche unter natürlichen Bedingungen wahrscheinlich noch weniger leicht erkennbar. Diese einseitige Erfahrung, so schließen die Autoren, könnte den Erfolg solcher Morphen beschränken und vielleicht sogar dazu beitragen, dass sie so selten sind. Die Fallstudie aus Arizona mit dem Titel White flowers finish last: pollen-foraging bumble bees show biased learning in a floral color polymorphism ist großartige Forschung - aber es sollten weitere Arbeiten folgen, um zu sehen, ob Blüten mit Farbpigmenten von vornherein einen Vorteil gegenüber hypochromen Blüten haben.